
一般財団法人環境イノベーション情報機構

2023.01.10
神戸大学内海域環境教育研究センター 特命教授 川井 浩史
川井 浩史(かわい ひろし)プロフィール

マングローブや藻場(もば)などの沿岸域の生態系は高い二酸化炭素(CO2)吸収能を持っており、地球規模の温暖化防止において、陸域の生態系に匹敵する大きな役割を果たしています。2009年には海で固定・貯留される炭素を「ブルーカーボン」と呼ぶことが提唱され、我が国においてもこの数年、この言葉を頻繁に耳にするようになりました。ここでは、「ブルーカーボン」のうち、日本の沿岸で特に重要な役割を果たしており、また将来の有望なCO2吸収・貯留対策として注目されている海藻や海草と、それらが作る藻場生態系の基本的な事柄について解説します。
20世紀以降、化石燃料の使用やセメント製造などによる人為起源のCO2(二酸化炭素)の放出が急激に増加しました。これらの大気中のCO2は、約30%が森林などの陸域の生態系で吸収されていますが、それとほぼ同等のCO2が海洋で吸収されています。これをふまえて、後述するように国連環境計画(UNEP)などによって、海洋で吸収・貯留される炭素を「ブルーカーボン」と呼ぶことが提唱されました。ちなみにこれにあわせて、最近では「グリーンカーボン」は陸域で吸収・貯留される炭素だけを示すようになりました。そして、これらのいずれによっても吸収しきれない約40%のCO2が大気中に残留し、温室効果ガスとして温暖化を引き起こしています。
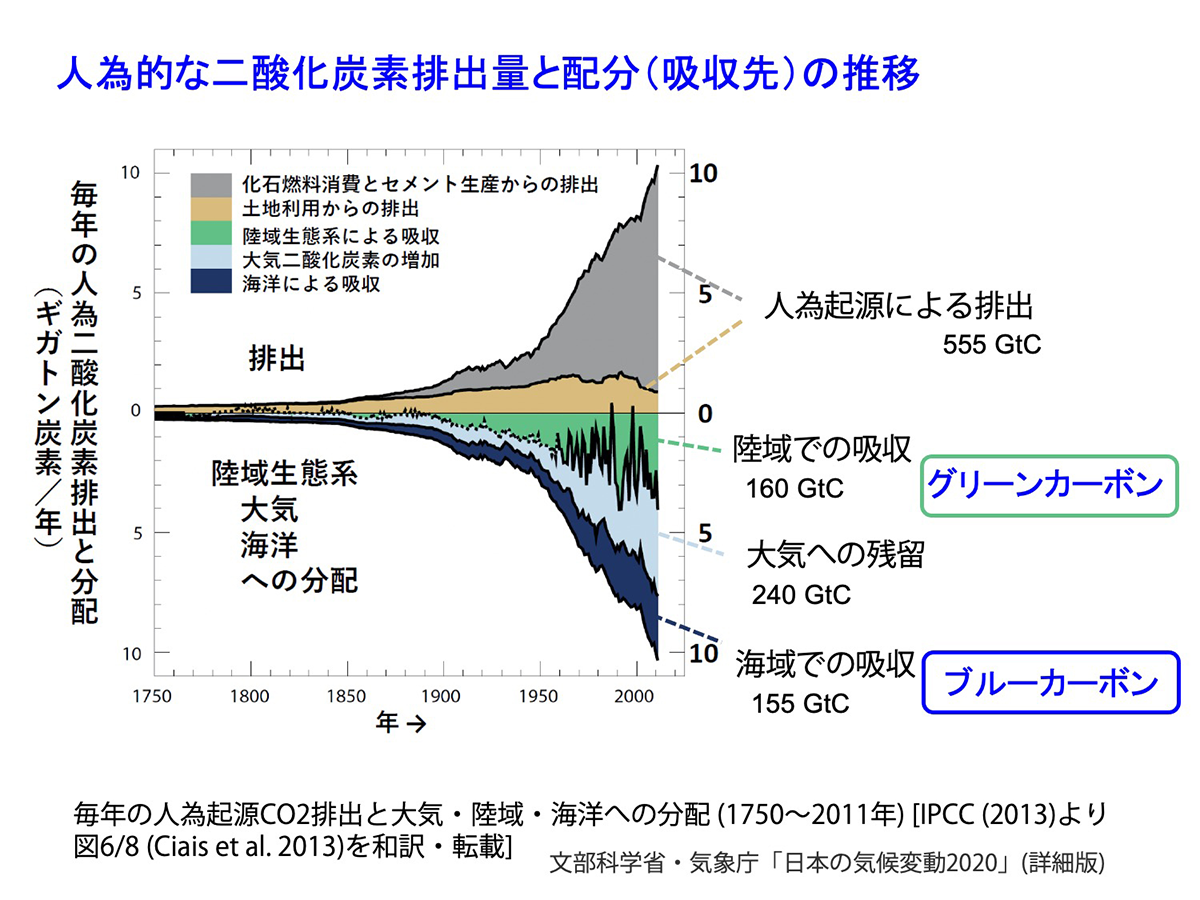
図1 人為的な二酸化炭素排出量と配分(吸収先)の推移 [画像クリックで拡大]
海でのCO2の吸収・貯留を担っているのは、陸域との接点にあるマングローブや塩性湿地の生態系のほか、植物プランクトン、海藻、海草などの海洋生態系を構成するさまざまな光合成生物です。しかし、森林の大規模な伐採や開発、火災、砂漠化などによって陸上生態系が劣化し、そのCO2吸収能が大きく低下しているのと同様、海洋生態系でもマングローブの伐採、藻場(もば)と呼ばれる、大形の海藻・海草が繁茂する沿岸域の生態系の急激な劣化・消失により、そのCO2吸収能の低下が深刻化しています。
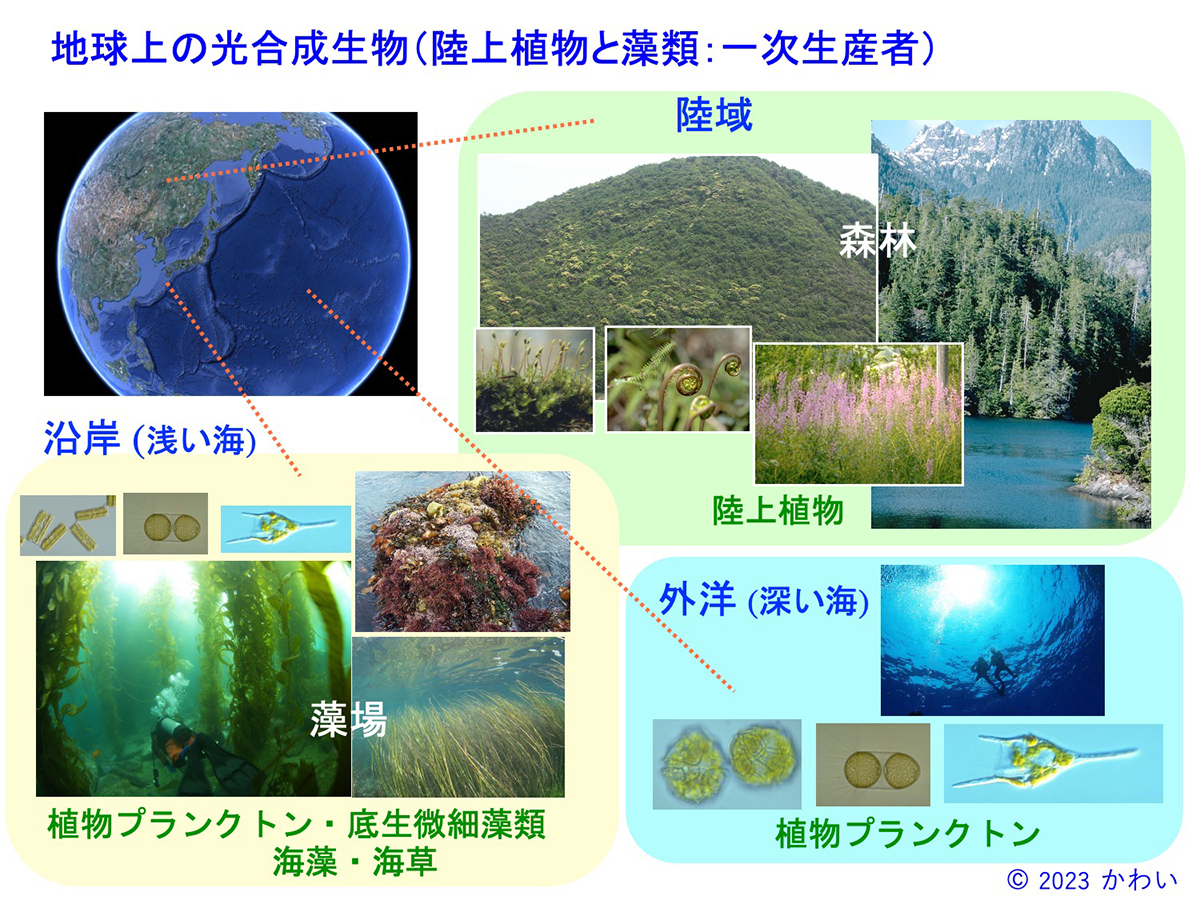
図2 地球上の光合成生物(陸上植物と藻場:一次生産者) [画像クリックで拡大]
地球表面の約3分の2は海ですが、その9割以上は水深200mより深く、海底には光合成に必要な量の光が届かないため、そこでは光合成によって生活する生物(いわゆる植物)は生存できません。このため、外洋の生態系は水面近くに留まることができる植物プランクトンが、主要な一次生産者となっています。しかしながら、外洋は光合成に必要なリンやチッ素などの栄養塩が少ないため、陸域から栄養塩が豊富に供給される沿岸域と比べると植物プランクトンの生物量は限られています。一方、海底まで充分な光が届き、栄養塩も豊富な沿岸の浅い海では、植物プランクトンに加え、海藻、海草などが生育しており、藻場(もば)と呼ばれる、陸上の森林に匹敵する立体的で、多様な生物が生活する生態系を構成しています。
海洋における生物量の90%近くはプランクトン、バクテリア、ウイルスなどの微細な生物が占めており、海洋の一次生産の95%以上は植物プランクトンが担っているとされています(Sogin et al. 2006; Suttle 2007)。毎年36.5GtのCO2が植物プランクトンによって吸収・固定されていると報告されていますが、そのうちどの程度が海底に有機炭素として貯留されるかについての推定値は、数%程度から10~20%と大きな幅があります(Gonzalez et al. 2008, Seiter et al. 2005)。
陸域生態系の一次生産者はほとんどが緑色植物(コケ、シダ、裸子植物、被子植物などの陸上植物)ですが、海域生態系はさまざまな系統群に属する「微細藻類」(植物プランクトンや底生微細藻)や「海藻」、「海草」など多様な光合成生物によって構成されています。これらのうち、「微細藻類」は生物量が非常に大きなものだけでもシアノバクテリア(らん藻)、珪藻、渦鞭毛藻、ハプト藻、ラフィド藻などがあり、そのほかにも多くの系統群が含まれています。一方、「海藻」は紅藻(アマノリ類、テングサ類など)、褐藻(モズク類、コンブ類、ホンダワラ類など)、緑藻(アオサ類、イチイズタ類など)の3つの系統的に異なる生物群の総称であり、これらは別々の系統の微細藻類が、独立して多細胞化、大形化して進化したものです。また「海草」は後述するように海で生活するようになった被子植物です。

図3 沿岸域の光合成生物 [画像クリックで拡大]
これらのさまざまな光合成生物はおおむね次のような道筋をたどって進化、多様化し、一次生産者として現在の地球上の生態系の基礎を担っています。すなわち、はじめ原核生物であるシアノバクテリアが、光合成により有機物を合成する能力を獲得しました。生命が誕生した頃の地球表面は嫌気的な環境でしたが、光合成反応では副産物として、水に含まれていた酸素(O2)が水中、または大気中に放出されます。このため、シアノバクテリアの大繁殖によって放出された酸素で地球表面は好気化し、その後、シアノバクテリアがアメーバのような従属栄養の真核生物と細胞内共生することで葉緑体となり、真核の藻類が誕生しました。これらの藻類が、「植物プランクトン」や「底生微細藻類」であり、その一部(緑色の藻類の系統)は淡水環境への適応を経て、陸上に進出しました。これが陸上植物の祖先であり、その後、コケ類、シダ類、裸子植物などを経て被子植物が出現し、これら植物の進化と並行してさまざまな動物や菌類が進化したことで、現在の陸上の生態系が成立しました。陸上植物は重力が大きく乾燥する地上の環境に適応する過程で、体を支え、土壌から水と栄養塩を吸収する構造である根を進化させました。
これらの被子植物のうち、海洋環境に進出したものがアマモなどの「海草」です。一方、海では陸上に進出することなく、海中の環境で定着、多細胞化、大形化するものがいくつかの系統であらわれ、それらが紅藻、褐藻、緑藻(アオサ藻)などの「海藻」に進化しました。
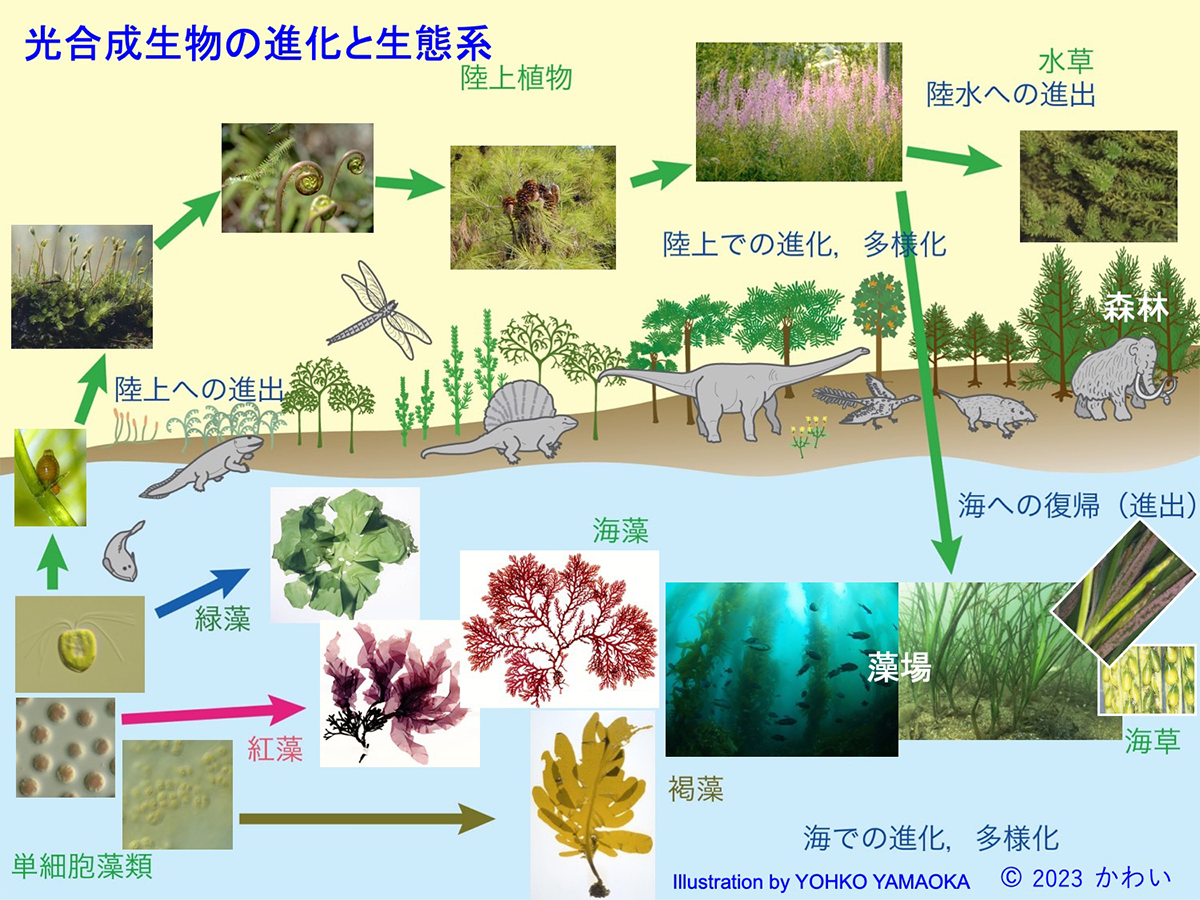
図4 光合成生物の進化と生態系 [画像クリックで拡大]
海藻のうち、褐藻の一部のものは10メートルを超えるきわめて大形の藻体を作り、それらが繁茂することで藻場(もば)と呼ばれる、陸上の森林に似た景観や機能を持った生態系が成立します。海藻は陸上植物と異なり、真の根を持たず、仮根と呼ばれる構造で岩や他の生物などに着生し、体の表面から栄養塩を吸収して成長します。また、ホンダワラ類やコンブ類の一部のものは、気胞と呼ばれる浮き袋を持っており、その大きな浮力で海底から立ち上がることで水面近くまで成長し、効率よく光合成を行っています。

図5 海藻藻場を作る大形褐藻類 [画像クリックで拡大]
海草は海に進出した被子植物で、海底に根を張って体を支えると共に、根から栄養塩を吸収して成長します。このうち、アマモ、コアマモなどは干潟などの砂泥質の海底を好んで生育し、アマモ場と呼ばれる藻場を作ります。アマモ場では密生する植物体の根元に泥などの懸濁物が堆積しやすく、そこに海草自体だけでなく海藻やプランクトンの死骸などが埋没し、貯留されることで面積当たり熱帯雨林の10倍を超える、炭素固定能を持つことが報告されています(Gacia et al. 2002, Nellemann et al. 2009)。一方、スガモやエビアマモなどは岩礁の岩の隙間などに根を張って生育し、スガモ場と呼ばれる藻場を作りますが、しばしば大形海藻と混生します。

図6 海草藻場を作る海草類 [画像クリックで拡大]
藻場はその生態系の主要な構成種によって、コンブ場(コンブ類、チガイソ類など)、ガラモ場(ホンダワラ類)、アラメ・カジメ場(アラメ、カジメ、クロメなど)、アマモ場(アマモ、コアマモなど)、スガモ場(スガモ、エビアマモなど)とさまざまな名称で呼ばれていますが、いずれの藻場も陸上の森林と同様の機能(生態系サービス)を持っています。すなわち、直接または間接的にさまざまな動物の餌となるほか、着生場所や産卵場所を提供し、また、ナーサリー、隠れ場となるほか、水質・底質の保持や波浪影響の低減、炭素の隔離(ブルーカーボン)などの役割を果たしています。

図7 藻場の生態系サービス [画像クリックで拡大]
陸上植物、植物プランクトン、海藻、海草などの光合成生物はいずれも、炭水化物、タンパク質、脂質などの有機物を合成する過程で、炭素(C)をCO2や炭酸水素イオンの状態で吸収し、その体の中に有機炭素として保持しています。しかし、これらの炭素は光合成生物自身が、あるいはそれらを餌として摂取した動物が、有機物を分解して生活のためのエネルギーを取り出すときにCO2として放出されます。また、これらの生物は死後、バクテリアや菌類によって分解され、再びCO2として放出され、それらを吸収する光合成生物に再利用される形で、生態系の中を循環しています。しかし、体を構成していた有機炭素は、死後全てが分解、放出されるわけではなく、その一部、たとえば根や土壌中に埋没した体の一部は分解されずにその場所で貯留されます。一方、植物プランクトンや海藻は、陸上植物と比べて寿命が短いものが多く、また一般に岩礁域に生育することから、藻体が土壌中に埋没することはまれであり、その有機炭素のほとんどが海底などで貯留されることなく循環していると考えられてきました。
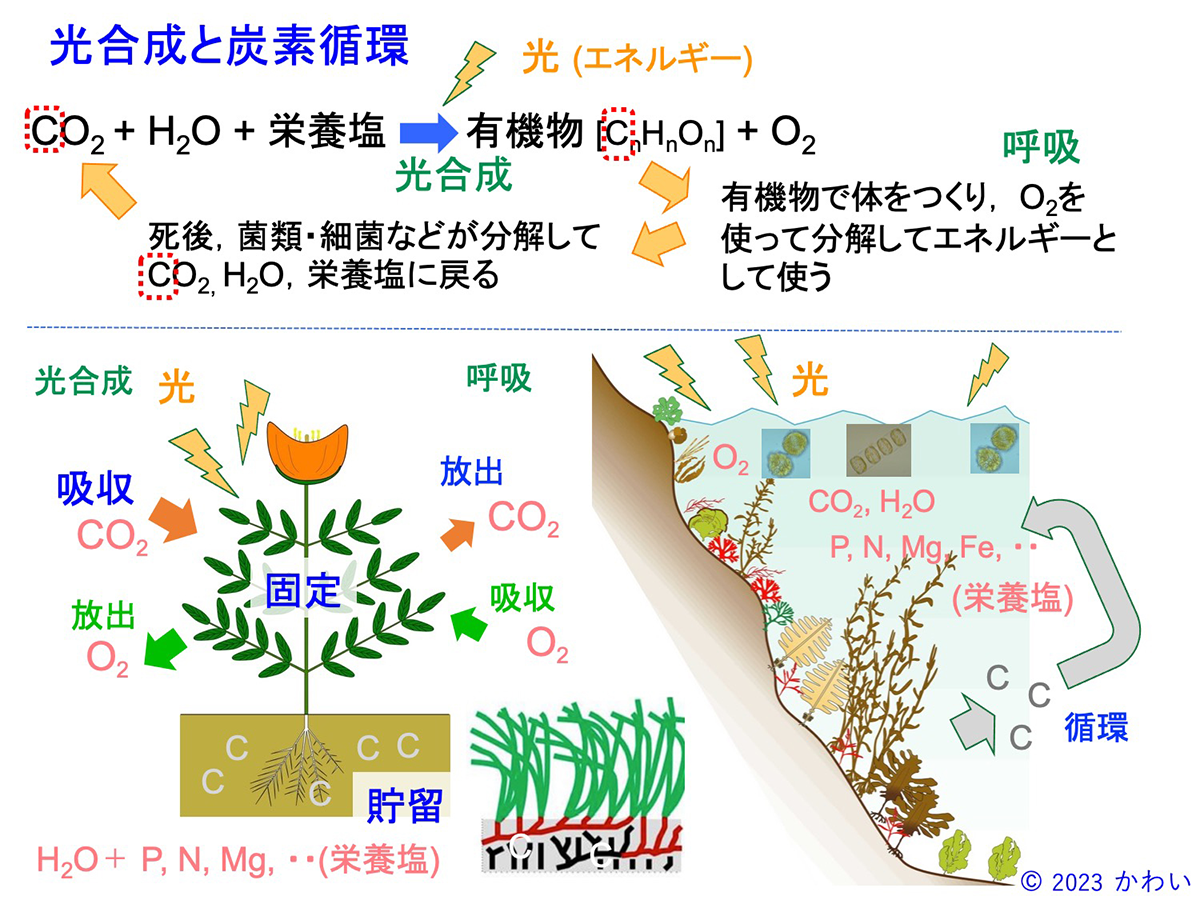
図8 光合成と炭素循環 [画像クリックで拡大]

図9 沿岸域各生態系の炭素固定・貯留速度の比較 [画像クリックで拡大]
国連環境計画(UNEP)、食料農業機関(FAO)、ユネスコ等によって2009年に発表された「ブルーカーボン」という報告書では、海域生態系のマングローブ、塩性湿地、海草藻場などが高いCO2固定能を持っており、その貯留量は陸上の生態系に匹敵することから、温暖化の防止に非常に重要であることが示されました。一方、植物プランクトンなどの微細藻類や海藻類は、寿命が短く、その炭素は死後速やかに水中に放出され、また海底にほとんど堆積しないと考えられることから、その炭素固定能は低いとされていました(Nellemann et al. 2009)。

図10 UNEP、FAO、ユネスコ等は2009年に「ブルーカーボン(Blue carbon)」という報告書を発表 [画像クリックで拡大]
これをふまえて、2013年に発表された気候変動に関する政府間パネル(IPCC)が出した湿地に関する「国別温室効果ガスインベントリーガイドライン補遺」では、各国が取りまとめることを義務づけられている温室効果ガスの排出・吸収量の一覧において、マングローブ、湿地、排水された土地、排水処理のために建設された土地における顕著な土地利用、管理の取り組みに対しては、インベントリー作成の項目が追加されましたが、海草藻場、海藻藻場については含められていません。

図11 2013年のIPCC国別温室効果ガスインベントリーガイドライン補遺に追加されたインベントリー作成の項目 [画像クリックで拡大]
S.V. Smithは1981年に海藻類、海草類の生物量は海洋の総生物量の3分の2に相当し、海藻草類の年間生産量は1×109t炭素と推定されることを報告しており、また、プランクトンが優占する海面が3.6×108km2であるのに対し、海藻草類が優占する海面は2×106km2にすぎず、海藻草類の生産効率は外洋の400倍に相当すると述べています。このため、海藻草類が高いCO2吸収能を持つことは、研究者の間では、知られていましたが、社会ではあまり注目されることはありませんでした。
その後、Krause-Jensen & Duarte(2016)は藻場生態系の炭素貯留に関する画期的な論文を発表しました。彼らはホンダワラ類やコンブ類などの大形褐藻の藻体が、ちぎれるなどして生育場所から流出し、深所の海底で堆積した海藻由来の有機炭素量を測定した結果から、数百メートル以深の無光層では光合成による炭素の再利用が起こらず、そのかなりの部分が貯留されることから、大形褐藻の藻場で固定された炭素の約10%が長期間にわたって海底で隔離されると推定しました。海藻藻場は一次生産量が非常に大きな生態系であり、またマングローブが発達しない温帯域で大きな面積を占めることから、この報告以降、炭素貯留源としての海藻藻場の役割の大きさや、温暖化対策としての活用の可能性が大きく取り上げられるようになりました。
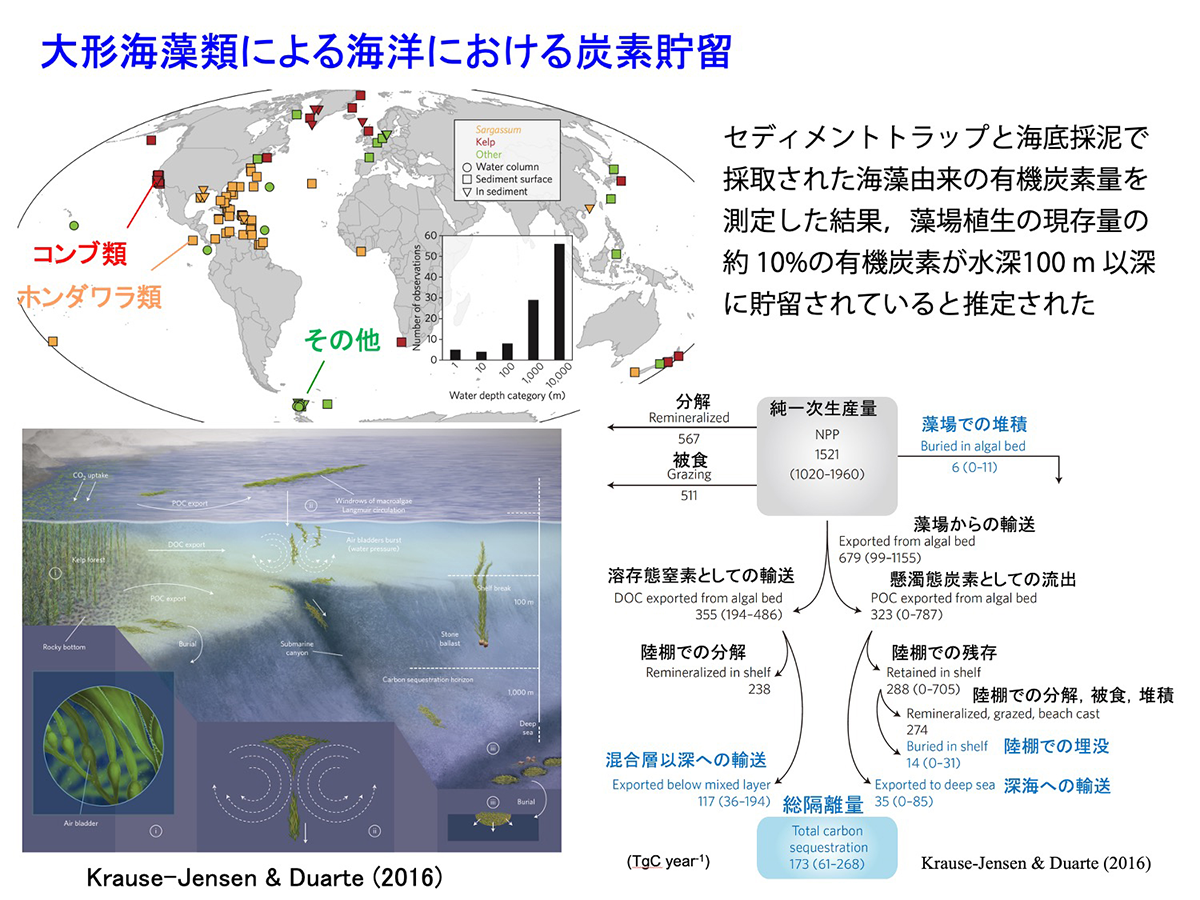
図12 大形海藻類による海洋における炭素貯留 [画像クリックで拡大]
これは、温帯域でのブルーカーボンの概念を示す模式図です。UNEP等による報告書で定義されたブルーカーボンは本来、マングローブ、塩性湿地などを含み、地球規模ではこれらの環境要素が重要な役割を果たしています。しかし、後述するように日本の沿岸ではこれらの植生は限られており、このため海藻、海草に焦点を当てた図としています。
CO2は水によく溶けますが、それでも通常は大気中のCO2濃度と、水中のCO2濃度はほぼ平衡状態にあり、植物プランクトンや海藻草類などの水中の光合成生物などが、吸収・固定した分だけ新たに大気から水中に溶け込むことになります。光合成生物によって固定された炭素は、その体の構成成分として保持されますが、死後、海底の土壌中に堆積した部分、または無光層より深い海底に沈降した部分が長期間にわたり貯留されることになります。また、海上で養殖されている海藻類や、植物プランクトンなどによって吸収されたCO2も、無光層より下に沈降した有機炭素の部分は貯留されます。ただ、外洋は栄養塩が少ないため、植物プランクトンの生物量は限られており、またかなりの部分は有光層で再利用され循環しています。
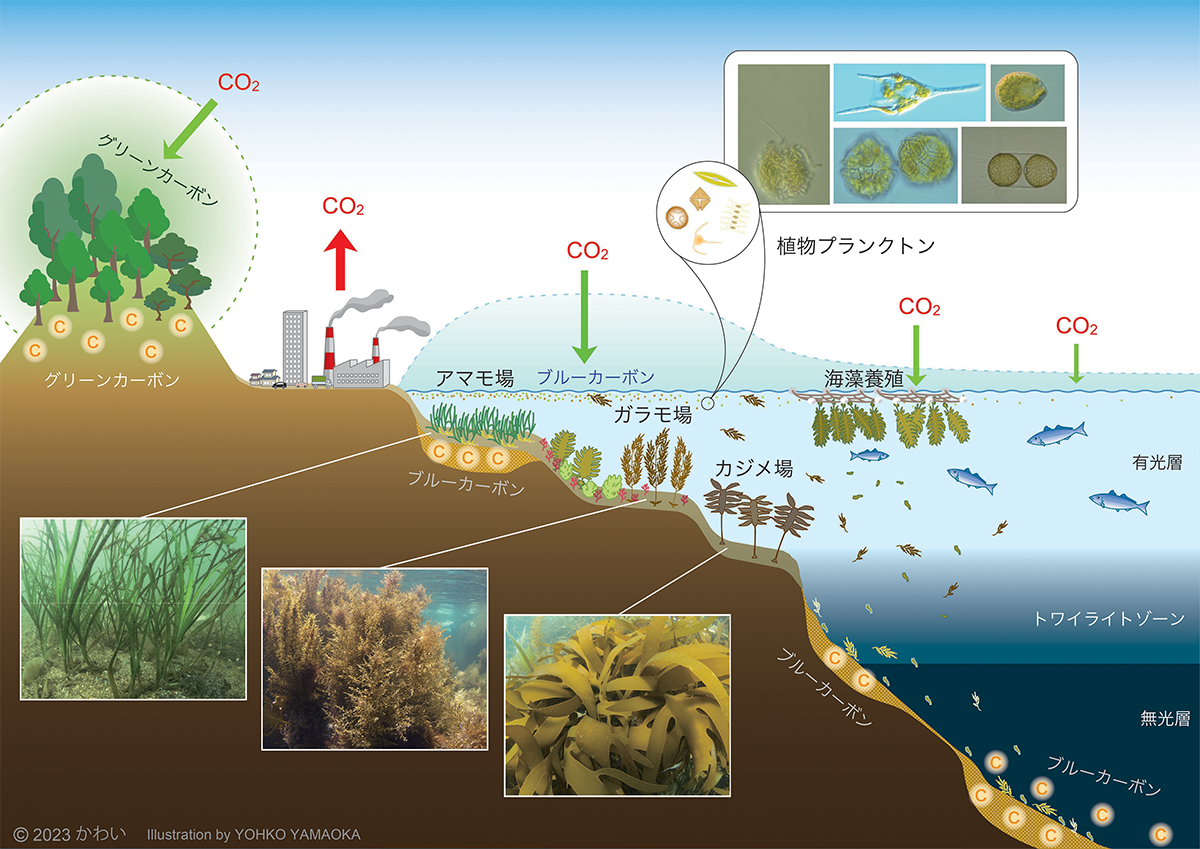
図13 温帯域でのブルーカーボンの概念を示す模式図 [画像クリックで拡大]
前述のUNEPほかによる2009年の報告書「ブルーカーボン」では、森林や海洋の持つCO2の吸収・貯留の機能を維持・拡大していくことの気候変動の緩和に向けての重要性が述べられており、その方策として陸域生態系を対象とした市場経済の仕組みを利用したクレジット化による支援と同様に、海洋生態系についても同様の制度を整備することが提案されています。しかし、前述したように現時点では国際的には藻場について、その炭素隔離・貯留量の評価、認証についての制度は整備されていません。
一方、国内では、地方公共団体や公的研究機関などが独自に藻場の創出・整備などの実績に対して、その評価、認証を行い、クレジット化する仕組みが作られています。

図14 国内におけるブルーカーボンの評価、認証、クレジット化の事例 [画像クリックで拡大]
日本は亜熱帯から冷帯に及ぶ長い海岸線を持ち、その多くは岩礁になっています。このため、衛星画像解析などを元に推定された海藻藻場(約1230km2)とスガモ場(約90km2)をあわせた面積は、アマモ場(約330km2)の3~4倍程度になっています。一方、「ブルーカーボン」の報告書(Nellemann et al. 2009)でその役割が強調されていたマングローブは、日本沿岸での面積は、約6~7km2ときわめて限られています。
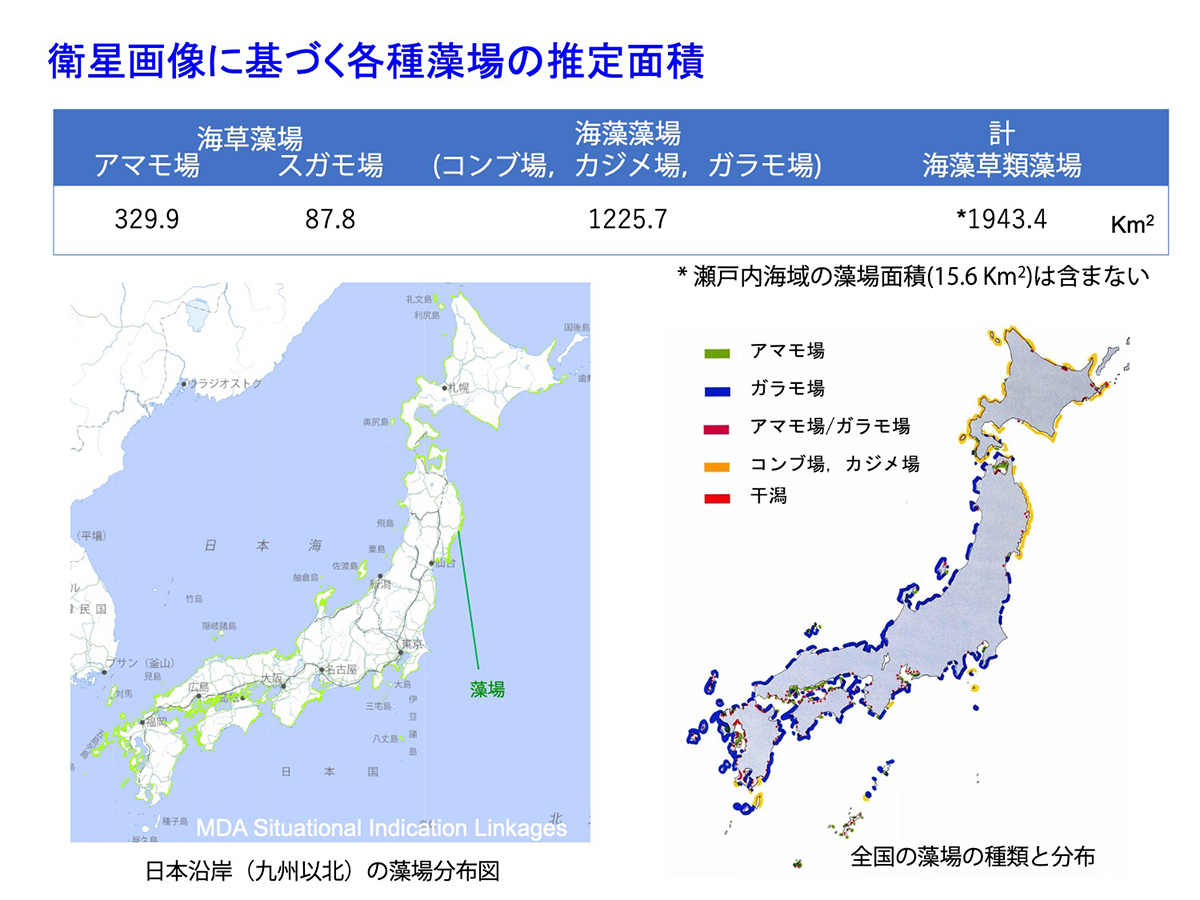
図15 衛星画像に基づく各種藻場の推定面積 [画像クリックで拡大]
海藻藻場を構成する大形褐藻類の種は多様であり、藻場の種類や海域の環境によって一次生産量は大きく異なる上、それらの種の特性(例えば浮力の違いによる外洋・深所などへの運ばれやすさ、死後の分解されやすさ、生育期間など)や周辺の海底地形・水深などによって、有機炭素の貯留効率も大きく異なると考えられます。しかし、これらの事柄についての研究はまだ非常に少なく、日本周辺の海域でどの程度の炭素貯留が起こっており、また今後、どの程度、炭素貯留源として活用可能なのかについては不明な部分が多く残されています。
富栄養化による透明度の低下や懸濁物の増加や、埋め立て・防波堤の設置などに伴う海水流動の低下、護岸工事による着生基質の単純化などにより、海藻が生育できる水深が浅くなり、また種の多様性が低下するほか、藻場を構成する大形の種類が生育できなくなります。この様な海域では、しばしば海底に沈降した有機物を多く含む堆積物によりヘドロが発生し、CO2の20~25倍の温暖化効果を持つとされるメタン(CH4)が発生するほか、季節によっては貧酸素化して硫化水素を生じ、好気性の生物の死滅を引き起こします。このような劣化した藻場生態系は、温室効果ガスの放出源となります。
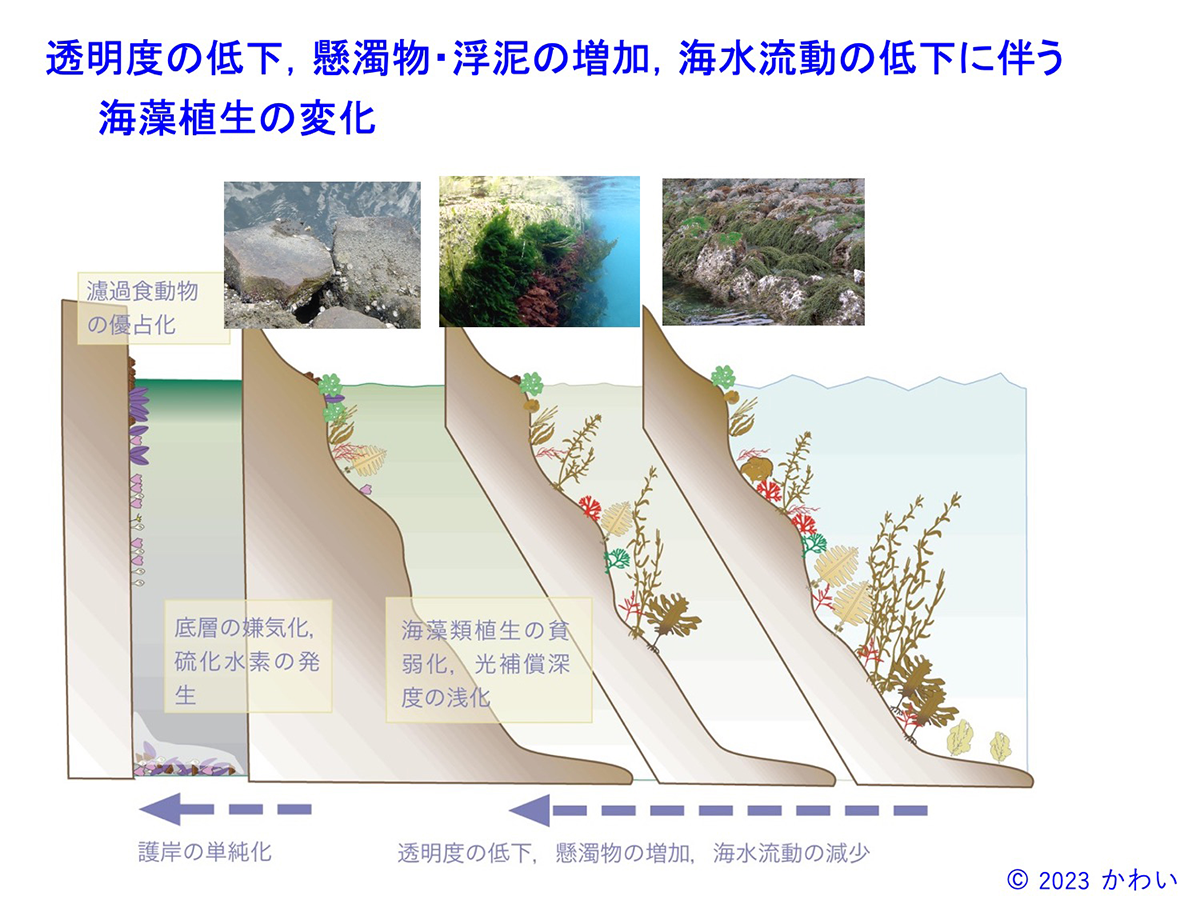
図16 透明度の低下、懸濁物・浮泥の増加、海水流動の低下に伴う海藻植生の変化 [画像クリックで拡大]
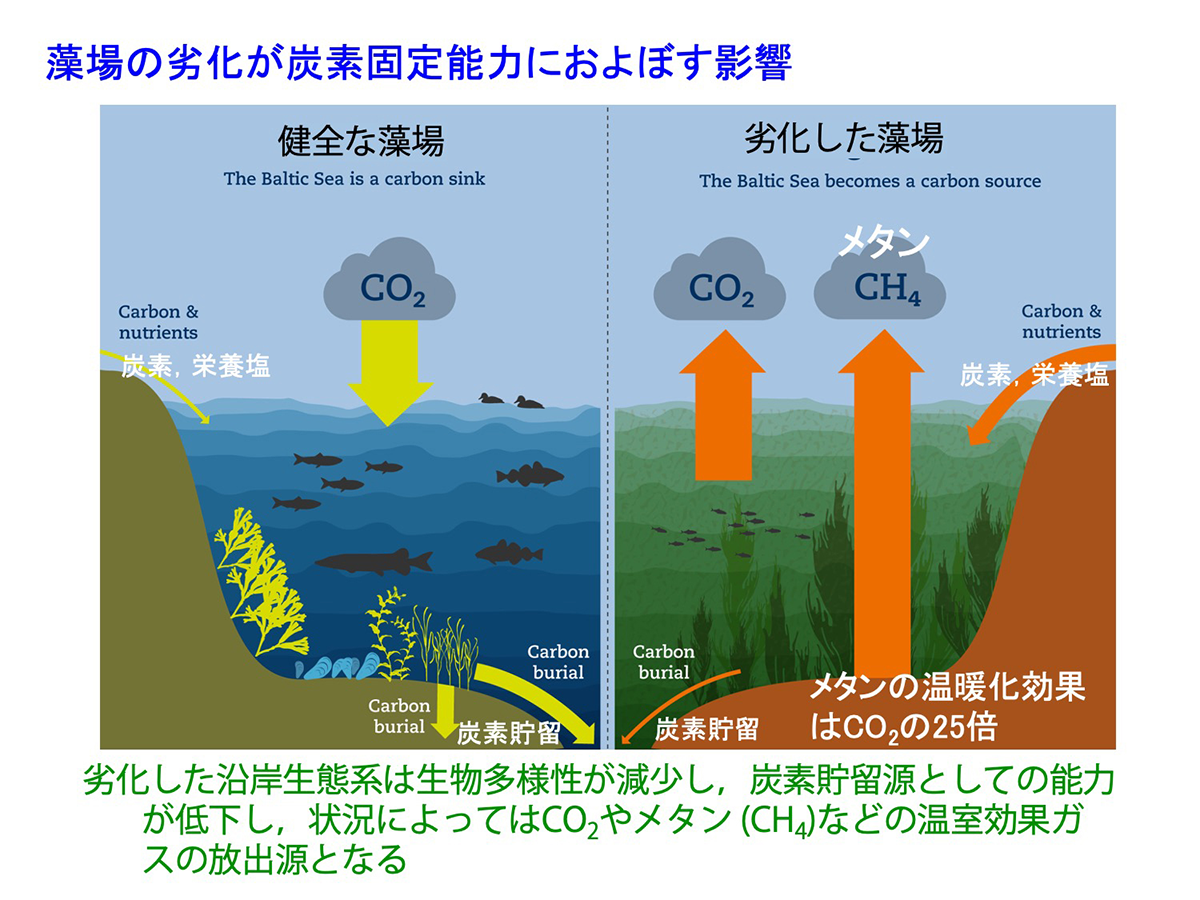
図17 藻場の劣化が炭素固定能力に及ぼす影響(出典 Illustration: Robert Kautsky/Azote (Stockholm University Baltic Sea Centre). Healthy coastal ecosystems are crucial to mitigate climate change. https://www.su.se/stockholm-university-baltic-sea-centre/policy-analysis/policy-briefs-and-fact-sheets/healthy-coastal-ecosystems-are-crucial-to-mitigate-climate-change-1.590756 ) [画像クリックで拡大]
日本沿岸の海藻草類の藻場は高度経済成長期の水質悪化、埋め立て、護岸工事などによって、大きく減少してきました。このうち、海草藻場が発達する場所である干潟の面積は全国的に過去数十年で40%程度減少しており、瀬戸内海ではアマモ場は70%近く減少しています。海藻藻場については、以前の藻場面積についての情報が乏しいため、充分な比較が行えませんが、アマモ場同様、大きな減少があったと考えられています。さらに、近年は、広い範囲でいわゆる「磯焼け」による藻場の衰退が報告されており、特に本州中部以西の太平洋沿岸では、温暖化または黒潮の大蛇行の影響による高水温によって草食魚類の接触圧が高まり、藻場の衰退・消失が進んでいます。
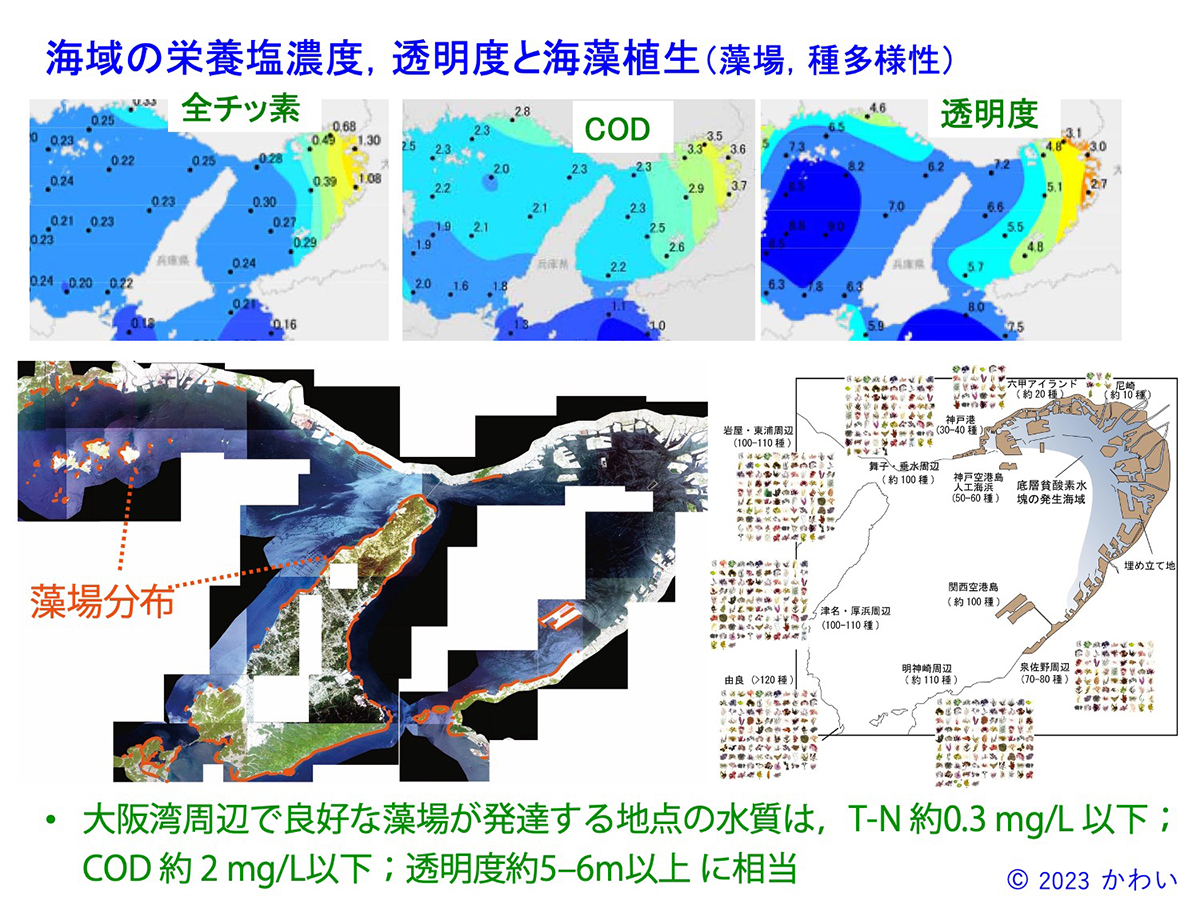
図18 海域の栄養塩濃度、透明度と海藻植生(藻場、種多様性) [画像クリックで拡大]
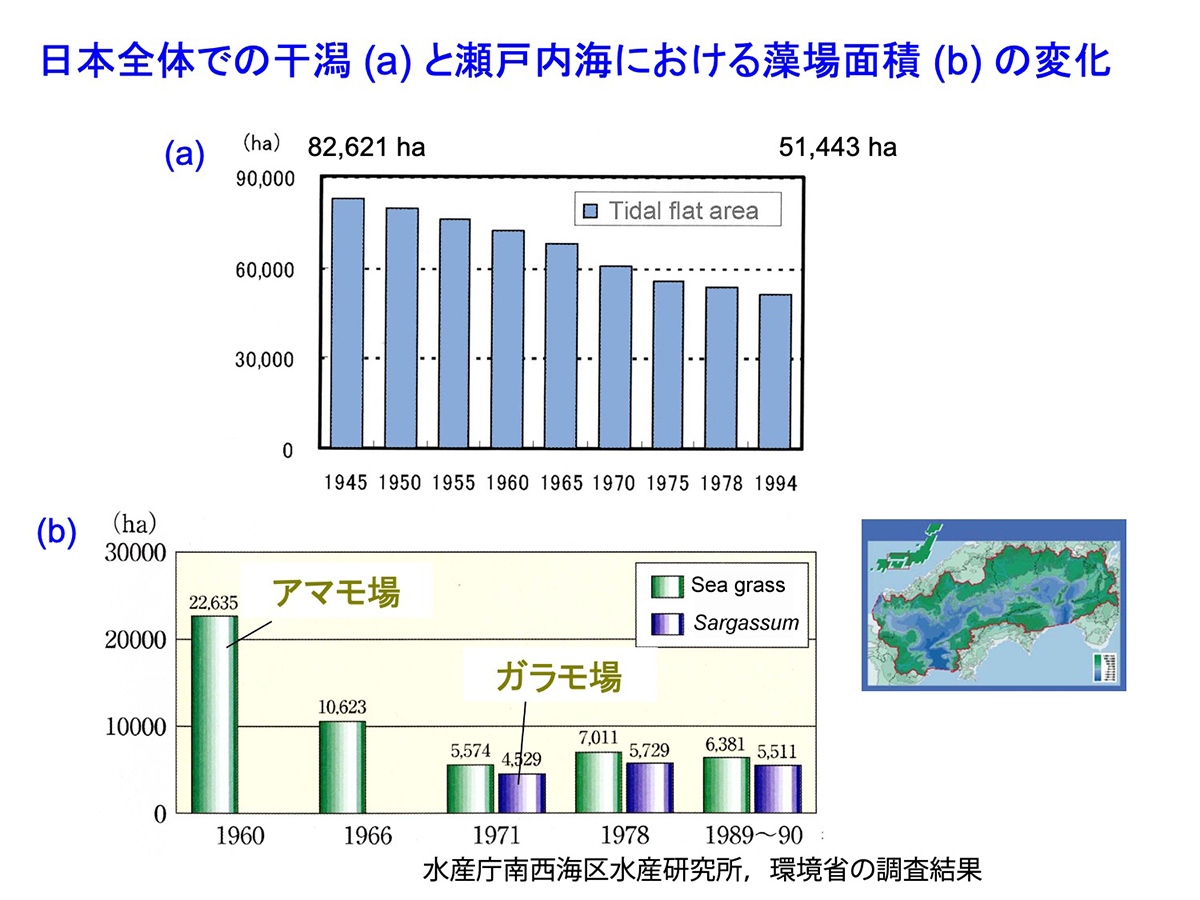
図19 日本全体での干潟(a)と瀬戸内海における藻場面積(b)の変化 [画像クリックで拡大]

図20 磯焼け [画像クリックで拡大]
一方、コンブ類、ワカメなどの大形褐藻やアマノリ類、オキナワモズクなどの海藻類が日本の広い範囲で養殖されています。これらの海藻養殖技術のほとんどは日本で開発され、普及したもので、日本は長い海藻養殖の歴史を持っています。この技術を活用して、海藻を活用したCO2貯留が各地で検討されています。
日本の沿岸では、すでにかなりの海面に漁業権が設定されており、また実際に養殖などに利用されています。しかし、世界的に見ると、東アジアに限っても中国、韓国、フィリピンなどでは日本よりはるかに多くの海藻が養殖によって生産されています。日本でもこれまでにあまり利用されていない沖合の海面、たとえば洋上風力発電施設が立地する海域などの利用を図ることで、まだ生産を増やす余地は充分あると考えられます。
この記事についてのご意見・ご感想をお寄せ下さい。今後の参考にさせていただきます。
なお、いただいたご意見は、氏名等を特定しない形で抜粋・紹介する場合もあります。あらかじめご了承下さい。
※掲載記事の内容や意見等はすべて執筆者個人に属し、EICネットまたは一般財団法人環境イノベーション情報機構の公式見解を示すものではありません。